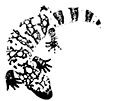
Riproduzione

-
In natura
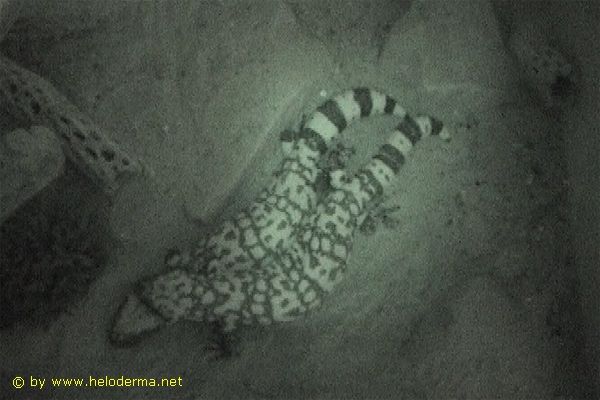
In un caso, un tentativo di accoppiamento è stato documentato "all'aria aperta", sebbene l'avvenimento di una copulazione non sia confermato (Rif. 79). Sinora nessuno è riuscito a osservare accoppiamenti di Heloderma al di fuori dei loro rifugi. Una sola deposizione di uova è stata documentata e resa pubblica (Rif. 20). Il momento esatto della schiusa è tuttora ignoto. Dato che i neonati (peso alla schiusa di circa 30-40 g) si osservano solo a partire dalla fine di aprile mentre la deposizione delle uova avviene in giugno, si può soltanto ipotizzare che i piccoli nascano in autunno entrando immediatamente in letargo oppure che l'embrione sviluppato rimanga all'interno dell'uovo anche nel periodo invernale per poi nascere in primavera (Rif. 21).
Le speculazioni in merito alla riproduzione di Heloderma suspectum si sono finite il 28 ottobre 2016 quando, durante alcuni lavori in un sobborgo settentrionale di Tucson, in Arizona, un escavatore ha aperto un nido che si trovava vicino al muro di un edificio, con all’interno cinque cuccioli di Mostri di Gila appena nati. L'intero ritrovamento è stato accuratamente documentato da Roger Repp (Rif. 67). Ora è dimostrato che Heloderma suspectum si schiude in autunno completamente sviluppato, per iniziare immediatamente il letargo sotterraneo.
-
In cattività
Gli Heloderma tenuti in cattività diventano sessualmente attivi soltanto al terzo anno di età. Gli esemplari maschi possono necessitare di un anno in più per raggiungere la maturità. I maschi devono raggiungere una lunghezza del tronco di ca. 22 cm, mentre le femmine di ca. 24 cm per essere sessualmente maturi (Rif. 22). Presupposto essenziale per un allevamento dai buoni esiti è un periodo di letargo di almeno tre mesi a una temperatura di circa 13°C. L'accoppiamento deve avere luogo entro il periodo di ovulazione della femmina, ossia quando sono maturi i follicoli. I maschi devono fornire spermatozoi completamente maturi e sviluppati.
Se la femmina ha una tana adatta per stabulare le uova, la deposizione procederá in modo ottimale se la femmina ha un sito adatto (Rif. 23). Ogni covata ha una media 4-5 uova, in casi eccezionali puó deporre sino a 7 uova. Sovente la femmina gravida, qualche giorno prima di deporre rifiuta il cibo. Una femmina puó deporre anche covate minute e sottili, accade qualora non sappiamo se sia gravida a causa delle sue dimensioni.
Deposte le uova la femmina si prende cura di ricoprirle con dune di sabbia – ora l’iter della deposizione puó dirsi terminato. Dopo di che la femmina rimarrá qualche giorno accanto alle uova col presupposto di accudirle e proteggerle.
Le uova devono essere trasferite al piú presto nell’Incubatore. Nel caso in cui l’umiditá dell’incubatrice sia inferiore al 98% Rh, dovete inserire nell’incubatrice due vaschette piene per metá d’acqua per far ricrescere l’umiditá. Inserite una vaschette nell’area inferiore dell’incubatrice mentre l’altra nella parte superiore. Dopodiché i valori d’umiditá dentro le caselle d’incubazione rientreranno vicini al 100% Rh.
Donna mostro guardia loro uova, recante




Il tempo di incubazione delle uova varia da 134 a 155 giorni (ca. 5 mesi!) a seconda della temperatura di incubazione (26-29°C). Durante l'incubazione, le Uova sane hanno l’odore boschivo e possono essere molto resistenti alle infezioni di funghi. L’embrione é connesso al tuorlo dal sacco vitellino, attraverso il cordone ombellicale. Da due a tre settimane prima della schiusa, le uova iniziano normalmente ad afflosciarsi e il piccolo necessita di ca. 48 ore per nascere. Durante l’incubazione il tuorlo verrá assorbito completamente, risucchiato nell’addome. (Rif. 24,25,26,27,28,29,30).
Le seguenti fotografie mostrano esempi del ciclo di riproduzione




 Video
Video
All'inizio della stagione degli accoppiamenti, i maschi mutano gli emipeni. Il maschio aggancia un arto posteriore attorno al ventre della femmina e vi introduce il proprio emipene. Lo sperma fuor uscito dal condotto termina in prossimitá della cloaca. Poi tramite i passanti irrigati siti sull'emipene, giunge all'interno della cloaca femminile. E' da considerarsi terminata la fertilizzazione, dal momento che lo sperma arriva alle ovaie attraverso gli ovidotti. (Ref.46). L'accoppiamento dura in media 2,5 ore.
Lo sperma secco rimasto all'esterno della cloaca femminile, indica che la copula é ben riuscita. Invece le tracce di sperma che trovate sulla cloaca del maschio, indicano un'improvvisa eiaculazione. Questo non é un indicatore di accoppiamento.
L'emipene non ha l'uretra. L'urina passa via la cloaca.




Hemipenis quando tentativo de copulation (Scanalatura sulla superfice dell 'emipene) 



Serie di esuvie di emipeni selezionati Eccesso di sperma poco dopo Copulation femminile cloaca
Sembra che le femmine preferiscano determinati esemplari maschi. Se la "chimica" è giusta l'accoppiamento sarà molto rapido. In caso contrario, potrebbero occorrere diverse ore di "sollecitazione" e la femmina potrebbe mordere il maschio.Osservando prima le specie da incoraggiare alla copula, si capiranno meglio i modelli piú attuali da introdurre. Le femmine verso i maschi nella loro condizione ambientale. In caso inverso i tempi del maschio saranno molto piú lunghi da marcare. Se vengono stabularli in ambienti nuovi corriamo il rischio che si scordino di copulare
Diagrammi di organi sessuali




A: Organi sessuali delle femmine
1) Uova pronte al deposito, 2) ovaie, 3) tubo di Falloppio, 4) bocca del tubo di Falloppiano nella cloaca, 5) colon, 6) ostitiumB: Organo sessuale del maschio
1) Ghiandole, 2) LATO ghiandola, 3) bocca del "tubo-sperma-urina" dentro la cloaca, 4) rene, 5) colonC: Emipene di Heloderma suspectum D: Emipene di Heloderma horridum alvarezi
A: Allo scoppio della parete delle cellule ovarica le uova mature fluttuaranno senza impedimenti nella cavità addominale. Poi si fondono all'insú verso l’apertura-fessura (ostians) dei tubo Falloppiano. Mentre vagano in direzione della cloaca sono fecondate da spermatozoi. (Rif. 54).
B: le ghiandole Neben allungate al fianco dei reni, costituiscono come una rete tessuta da canali sottili. I tubi di fuor-uscita formano delle vie spermatiche ai reni, seguendo i bordi centrali della parete. Prima dell’ingresso in cloaca si fonderanno col condotto orinatoio per poi entrare in cloaca attraverso il tubo (Rif. 54).
C, D: Gli emipeni sono in delle sacche ai lati destro e sinistro, sotto l’inizia della coda. Sono costituiti da cavi tubici, che possono trasformarsi dentro e fuori. A seconda delle specie ogni maschio deve espellere uno od entrambi gli emipeni per copulare. Le scanalature sulla superficie degli emipeni, faciliteranno l’irrigazione ed il trasporto degli spermatozoi nella cloaca femminile. La realizzazione di un emipene si determina per specifico in ogni specie.
Dimostrazione del ciclo completo di riproduzione con incluso: copula, guscio pre-uovo, deposizione e la schiusa dei cuccioli








GRAFICO SULLO SVILUPPO EMBRIONALE DELLE LUCERTOLE


A: Stato dell'uovo al momento del deposito
) Strato di minerali, 2) blastoderma, 3) membrana dell’uovo (sierosa), 4) tuorloB: Embrione molto giovane
1) Embrione, 2) amnio, 3) allantoide, 4) tuorlo, 5) sierosa, 6) sacco del tuorlo, D ) celoma extra-embrionaleC: Embrione giovane
1) impianto dell'embrione, 2) faglia amniotica, 3) allantoide, 4) tuorlo, 5) sierosa, 6) colonB: l'embrione s’è staccato un poco dal tuorlo. Colon e sacco vitellino sono collegati tra loro. Il tuorlo non è ancora circondato completamente dall\’endoderma. Sono in aumento le pieghe amniotiche davanti e dietro il corpo embrionale. La loro parete esterna si tramuta in corion (sierosa). È sostenuto da uno strato mesodermico, sito direttamente sotto la pelle delle sue membrane.
C: Le membrane embrionali sono completamente sviluppati ed il tuorlo è inserito del tutto all'interno del sacco vitellino. L’amnio é costituito da un sottile strato di ectoderma, coperto dal mesoderma. L’amnio si sviluppa in un sacco pieno di liquido. Quivi l’embrione galleggia come fosse in un piccolo stagno. La membrana corio-allantoidea serve da “placenta primitiva” garante degli scambi di sostanze metaboliche (prodotti). La cava dell’allantoide funge da deposito per azoto contenente prodotti (es.: acido urico) dell’embrione. Quando emergono i cuccioli dalla schiusa: l’amnio, il corion ed il corio-allantoide si trattengono dentro “l’Uovo”.
Lo squame della lucertola si sviluppa in tardo stato di crescita embrionale: in un primo tempo lo squame corporeo, in seguito proteggendo lo squame cervicale. I pigmenti appaiono prima negli occhi poi attorno l’organo parientale ed infine sulla pelle (Rif. 54).
SVILUPPO dell'EMBRIONE - COVA




Surface of egg with crystals Substrato senza uova di Heloderma in incubate Speratura di uova fertile, dopo la deposizione
Video
Uovo dopo 4 mesi di incubazione: nota bene la prominenza delle vene e la forma embrionale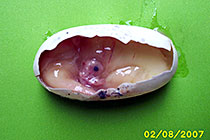


Embrione di circa 3 settimane di etá Embrione di circa 6 settimane di etá Documentazione grafica del battito cardiaco dell’embrione (Avitronics, Buddy Digital Egg Monitor) 
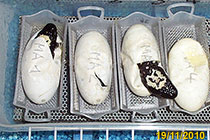

Singolo dente d'uovo di Heloderma Uova graffiate dal dente d'uovo Poco dopo la schiusa: addome voluminoso col tuorlo d'uovo
Quando un ovocita viene fecondato dall'Heloderma, entro 6 settimane si svilupperá attorno ad esso un guscio d'uovo morbido. Con la speratura subito dopo la deposizione, si rilevano le uova col blastoderma. Il blastoderma inizia a muoversi liberamente dentro l'uovo, 12 ore dopo la deposizione. Spostandosi verso la parete superiore della calotte (osservazione personale). La parete del guscio, una volta essiccata all'esterno, diventa permeabile all'aria e l'uovo può "respirare". La membrana vitellina si abbassa sul fondo dell'uovo: a essa è collegato l'embrione, che galleggia sopra al "plasma". L'embrione galleggia al di sopra del sacco vitellino attaccandosi al muro superior del guscio d'uovo adesso permeato d'ossigeno. Girare l'uovo ora distruggerebbe l'embrione in crescita – come accade coi serpenti ovipari. In questo momento il tuorlo d'uovo distrugge l'embrione in crescita - come nei serpenti ovipari, La mancanza di stringhe-di-GRAGNOLA sono responsabili della morte dell'embrione? (Ref. 45, 50, 54). D'ora in poi si sviluppa una rete differenziata di vene per rifornire l'embrione di ció che gli necessita.
Occorrono 5 mesi per crescere un embrione a circa 27 gradi. Il chiara d'uovo è sorgente di proteine e di sali minerali. Il tuorlo scende sul fondo d'uovo trattenuto dal sacco vitellino. Prima che il cucciolo lasci l'uovo, il sacco collegato al fegato si deve retrarre totalmente. Di modo che il tuorlo sia usato come primo pasto.
Alla fine quando l'uovo si svilupperá, spunterá un dente superiore in bocca della schiusa (praemaxilla). Quel dente gli consentirá di uscire dal guscio incidendo l'uovo. Costí é un dente vero e proprio, che perderá subito dopo la schiusa (Ref. 43, 46).
Differenza di dimensioni tra due piccoli e due esemplari di un anno. Alla schiusa il peso medio è di 35 grammi e dopo un anno è di circa 380 grammi in media.


-
Cicla di riproduzione
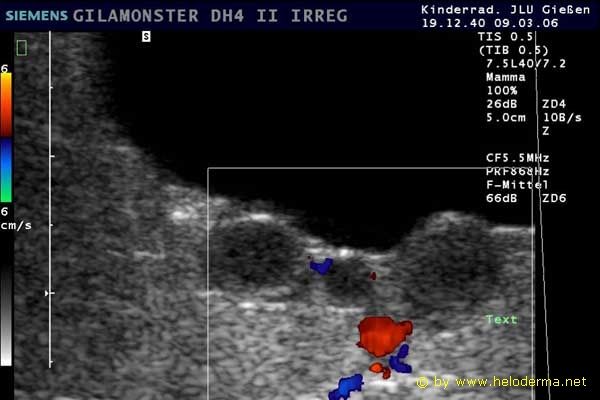
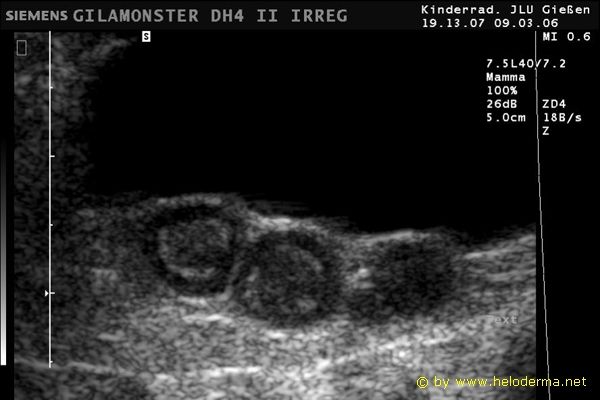
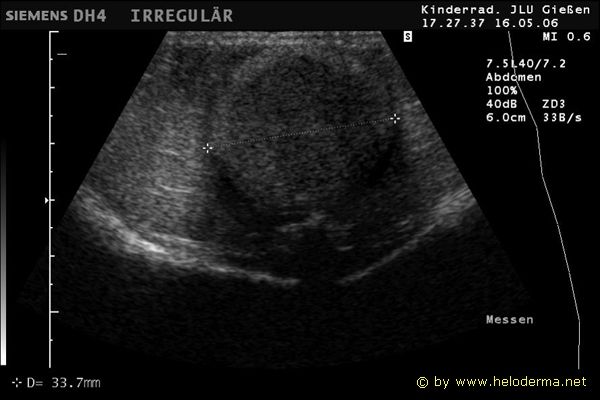
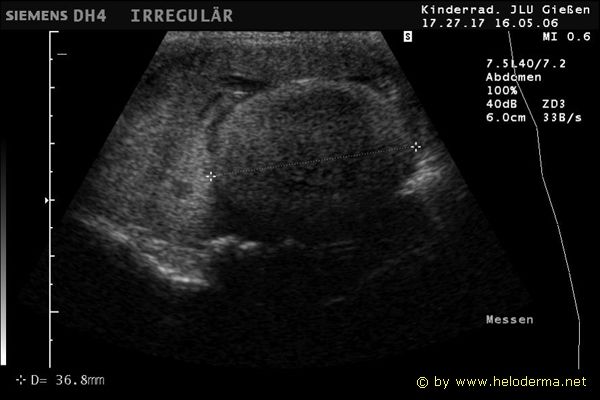
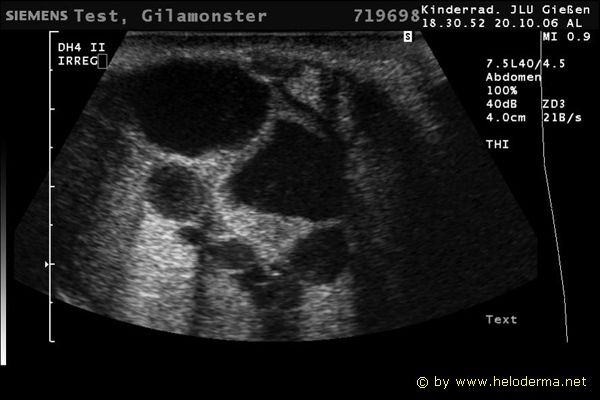
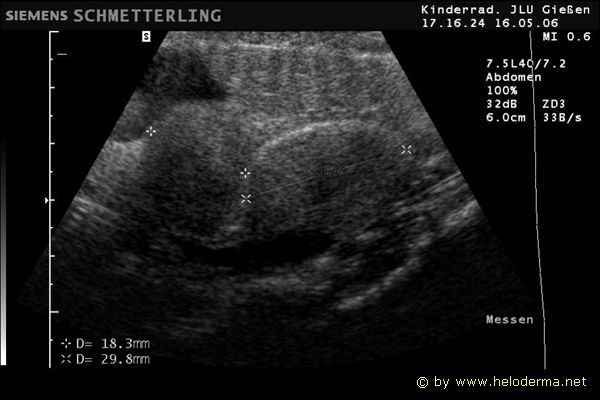
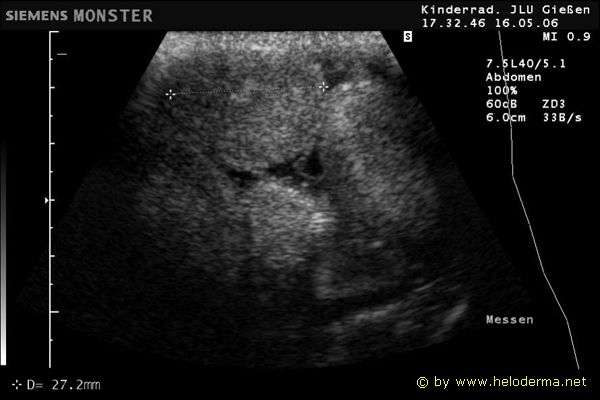
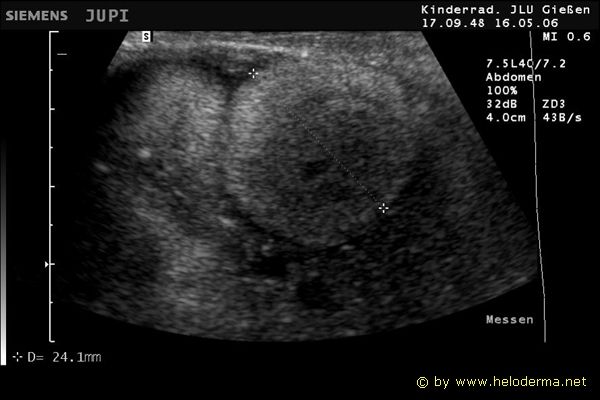
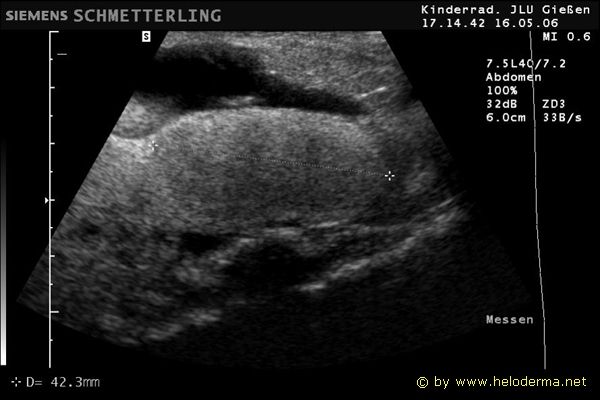
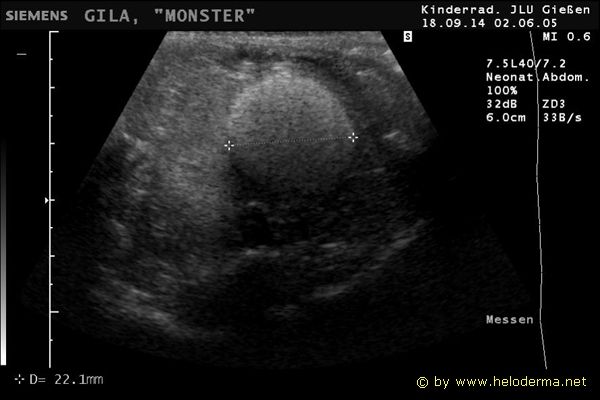
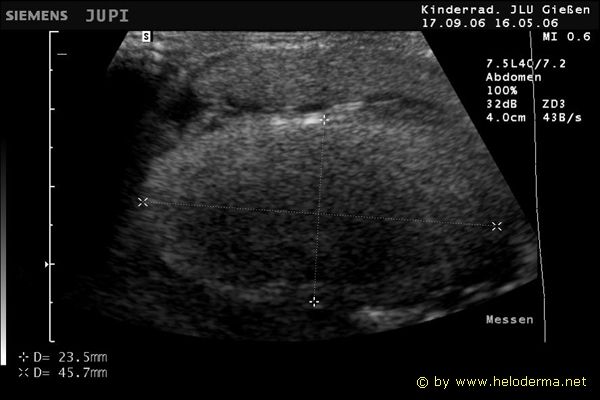
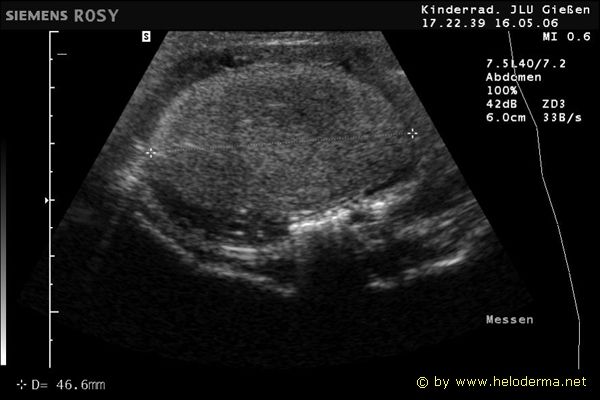
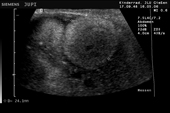
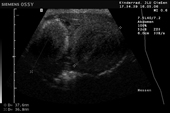
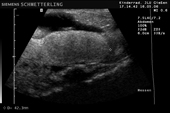
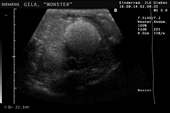
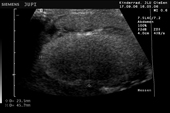
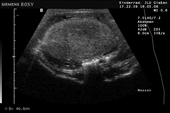
Controllo del ciclo di riproduzione mediante ecografia e raggi X.
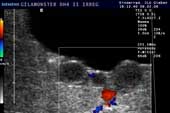
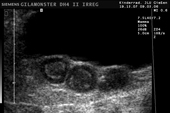
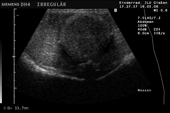
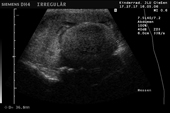
9.3.06 Follicoli sviluppat 9.3.06 Follicoli sviluppat 16.5.06 Uovo in fase di sviluppo 16.5.06 Uovo in fase di sviluppo 
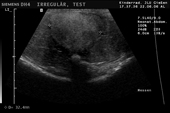
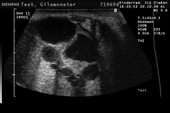

Inizio giugno 2006 22/06/06
L'uovo viene riassorbito?20/10/06
Appaiono nuovi follicoli per il ciclo a venire20/10/06
I raggi X non rilevano alcun uovo*
* Nota bene l’abbondanza di osteoderma, in piccoli punti bianchi sull’epidermide
Raffigurazione e misurazione di uova a ca. 3 settimane prima della prevista deposizione.
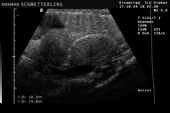
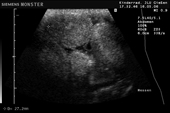
-
Combattimento
Il combattimento negli Heloderma è una lotta ritualizzata tra due esemplari maschi durante il periodo del corteggiamento e dell'accoppiamento. Il maschio dominante viene spesso preferito dalla femmina per l'accoppiamento (Rif. 31).
Esempi di un combattimento tra due maschi (molte foto sono abbinate a videoanimazioni).



 Video
Video 


 Video con il suon
Video con il suon
Video con il suon
Video con il suon
Video con il suon
Video con il suon
Video con il suon
Video con il suon
Dalla sequenza delle immagini si nota che la femmina nutre interesse e cerca il maschio.
Alcune sequenze del combattimento ricordano le posizioni di un corteggiamento maschio-femmina. L'osservatore può quindi confondersi tra il corteggiamento e il combattimento.
-
Corteggiamento
Per mostrare la differenza tra combattimento e corteggiamento sono state portate come esempio le seguenti videoanimazioni.

 Video
Video